

Mechanisms of resistance to targeted therapies
Composition de l'équipe
Directeur de Recherche
Chargé(e) de Recherche
Ingénieur
Doctorant(e)
Technicien(ne) Supérieur(e)
Activities
{kind=link}
{kind=link}
{kind=link}
{kind=link}
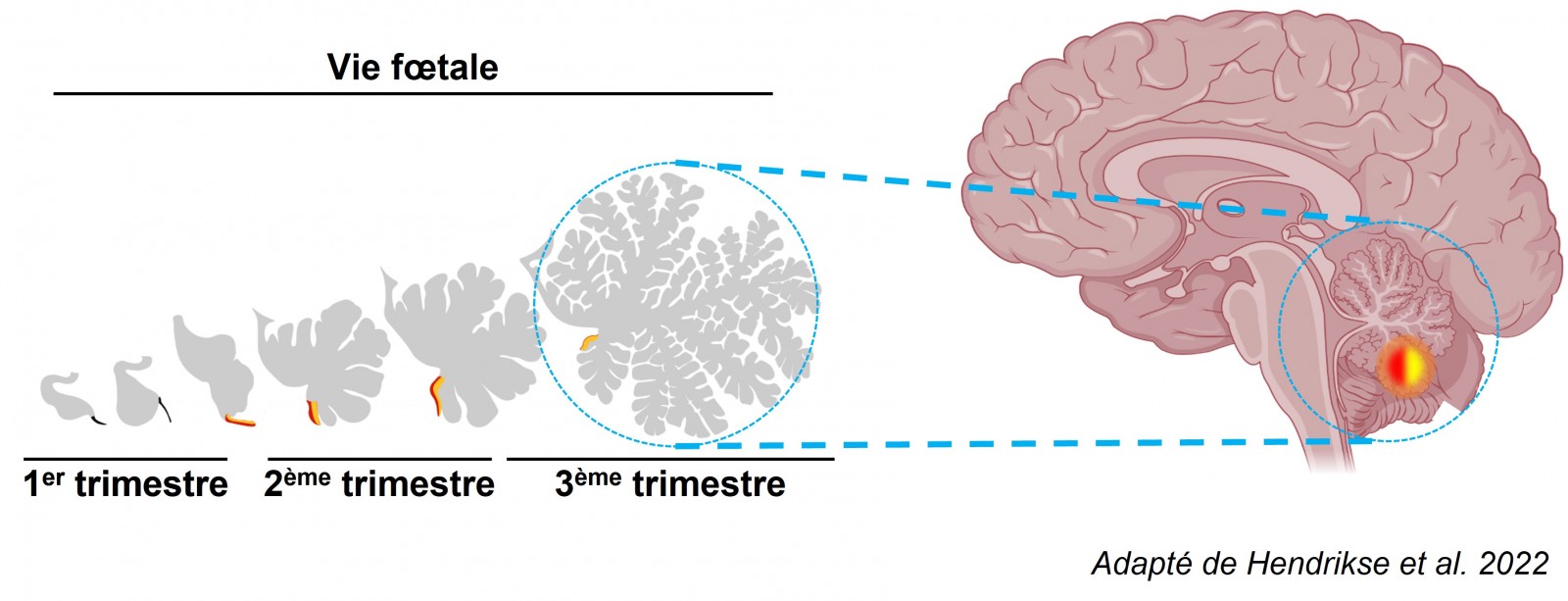
Notre équipe s’intéresse aux tumeurs du cerveau de haut grade. Ces tumeurs présentent la particularité d’être composées majoritairement de cellules dites peu différenciées, aux caractéristiques proches de celles des cellules embryonnaires du système nerveux en cours de développement. Cette observation a été rapportée dans plus de la moitié des tumeurs pédiatriques du cerveau, telles que les médulloblastomes, mais également dans les glioblastomes, qui peuvent toucher des adultes comme des enfants 1-4. La chimio- et la radio-thérapie constituent, avec la chirurgie d’exérèse lorsqu’elle est possible, les modalités thérapeutiques principales de ces cancers. Cependant, un nombre non négligeable de ces tumeurs (de 30% des médulloblastomes et 50% des épendymomes pédiatriques jusqu’à 90% des glioblastomes) présente une réponse incomplète à ces thérapies, entraînant des récurrences tumorales aux conséquences fatales4-6.
Une recherche collaborative
L’équipe Cellules Souches et Tumeurs du Cerveau bénéficie d’un réseau grandissant de collaborations nationales et internationales (Lurie Children’s Hospital, Chicago, USA ; Institut Gustave Roussy, Paris, France ; Laboratoire Radiopharmaceutiques Biocliniques, Grenoble, France, Institu de Biochimie et Génétique Cellulaires, Bordeaux…). L’équipe est également en étroite collaboration avec les équipes du Service de Médecine Nucléaire (dirigé par le Professeur Marc Faraggi) et du Service de Radiothérapie (dirigé par le Dr Cécile Ortholan-Nègre) du Centre Hospitalier Princesse Grace de Monaco. Notre équipe fait aussi partie du réseau de recherche en oncologie pédiatrique REACT4KIDS (REsearchers in oncology ACTing for kids).
Nos projets bénéficient d’un important soutien financier du Gouvernement Princier de Monaco comme d’associations telles que la Fondation Flavien, le GEMLUC (Groupement des Entreprise Monégasques de Lutte contre le Cancer) ou la Fondation Aleksander Savchuk.
Identité embryonnaire/souche des cellules de cancers du cerveau de haut grade
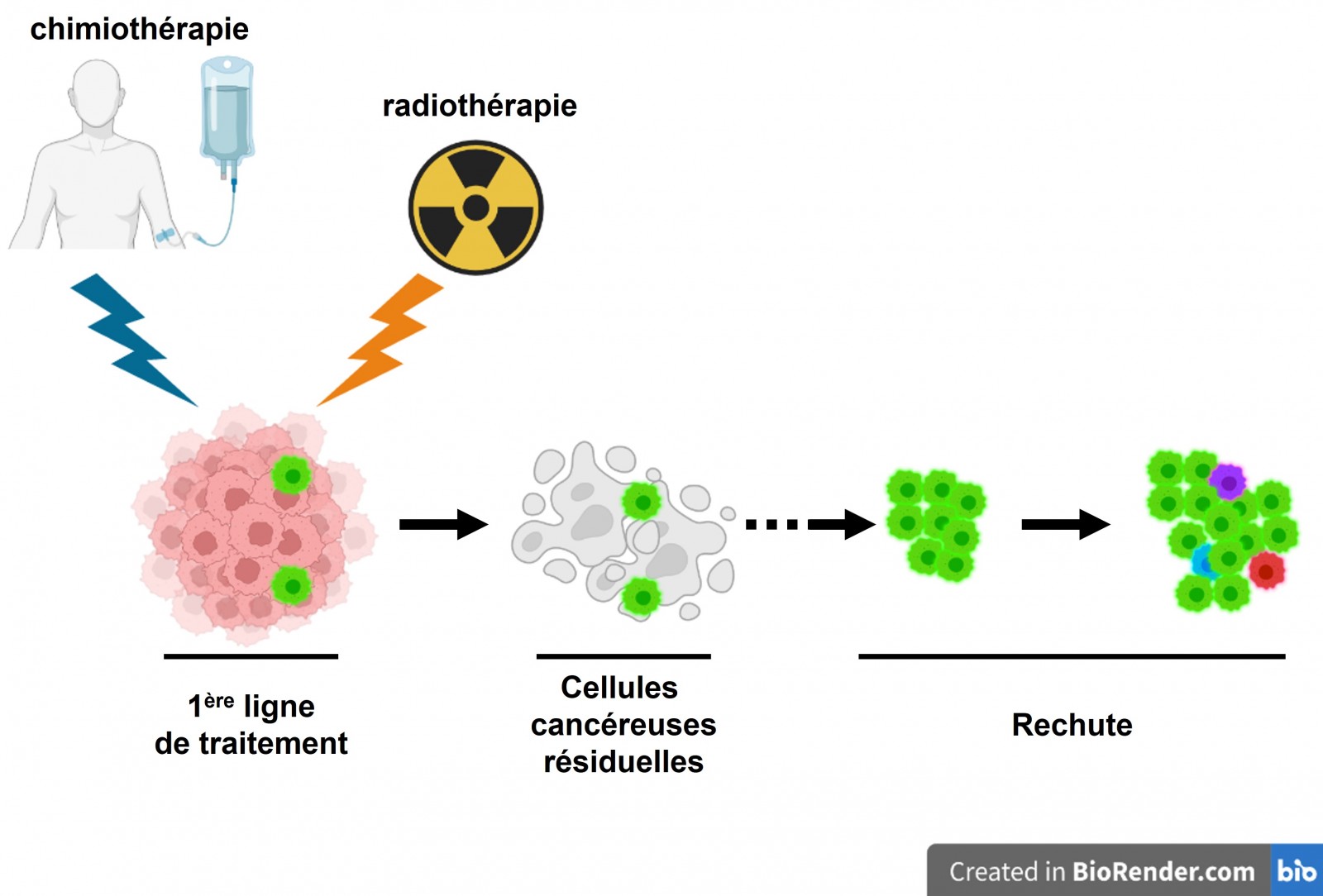
Un consensus scientifique est dégagé autour de l’idée que les rechutes de tumeurs cérébrales résultent de phénomènes d’adaptation ou de résistance naturelle aux traitements génotoxiques de certaines sous-populations cellulaires cancéreuses. Les données moléculaires cliniques et expérimentales concernant ces populations persistantes pointent en direction d’une identité souche et d’une quiescence des cellules les constituant1,7-9. Cependant, notre compréhension de la connexion qui semble exister entre les phénomènes d’initiation des tumeurs cérébrales, l’identité embryonnaire neurale des cellules les constituant et leur capacité à contourner les traitements génotoxiques reste extrêmement restreinte. Nos recherches consistent à interroger ces liens au travers d’approches aussi bien fondamentales que translationnelles. Cette démarche nous permet de nous placer en position de modéliser les propriétés particulières des tumeurs du cerveau à cellules souches et d’évaluer l’efficacité de modalités thérapeutiques de la manière la plus pertinente possible.
I) Projets de recherche fondamentale
I/1) Neurogenèse et tumeurs embryonnaires du cerveau (EmbryOnco)
La compréhension des mécanismes d’initiation des tumeurs cérébrales est au cœur du travail de recherche fondamental de l’équipe. Les données génétiques récemment obtenues par le séquençage massif de cancers pédiatriques du cerveau et la comparaison avec les tissus embryonnaires d’origine suggèrent que ces cancers trouvent leur origine pendant le développement embryonnaire. En effet, les cellules tumorales constitutives de nombreux cancers pédiatriques du cerveau sont tout à fait comparables avec des cellules embryonnaires neurales normales. Le projet EmbryOnco consiste à comprendre les événements à l’origine de la transformation de cellules souches et/ou embryonnaires du cerveau en cellules cancéreuses.
Au cours du développement embryonnaire normal, les cellules qui génèreront le système nerveux central prolifèrent et acquièrent graduellement une identité spécifique (neurones, astrocytes, oligodendrocytes…). Ce processus, dit de spécification ou de différenciation cellulaire, se conclu par un arrêt de la prolifération et l’acquisition d’une identité terminale stable. Nous avons fait l’hypothèse qu’un défaut de différenciation ou un « verrouillage » des cellules dans un état embryonnaire peut se solder par une prolifération anormale de ces cellules. Nous avons testé cette hypothèse en exprimant un gène responsable du maintien de l’état souche dans des cellules progénitrices neurales. Cette modification génétique a permis de rendre ces cellules cancéreuses et de former des tumeurs. Les analyses histopathologiques de ces tumeurs « artificielles » réalisées par le Pr. Fanny Burel-Vandenbos, du service d’anatomopathologie du CHU de Nice montrent qu’elle présentent des propriétés anatomopathologiques tout à fait comparables avec celles constatées chez les patients atteints de tumeurs embryonnaires du cerveau.
La validation de cette approche apportera des réponses à la question fondamentale des mécanismes conduisant à l’émergence de la tumeur et dotera notre laboratoire d’un système d’étude totalement original et pertinent.
I/2) L’anémone de mer Nematostella Vectensis comme modèle d’étude des cancers du système nerveux (Nematonco : in vivo investigation of embryonal neural oncogenesis in Nematostella vectensis)
Dans une démarche de développement de modèles d’étude novateurs, notre équipe développe un projet en collaboration avec celle d’Eric Röttinger (Université Côte d’Azur). Ce projet vise à utiliser l’anémone de mer N. vectensis pour mieux comprendre les mécanismes conduisant à l’émergence de cancers embryonnaires.
Les anémones de mer sont des invertébrés qui appartiennent, comme les coraux et les méduses, à la famille des cnidaires. Ces animaux sont dotés d’un système nerveux dit « diffus », constitué d’un réseau de neurones isolés sans ganglions nerveux - et encore moins de cerveau. Le très haut degré de similarité entre la neurogenèse des cnidaires et celle des vertébrés nous laissent présager que les découvertes réalisées sur ce modèle seront directement transposables aux mammifères. Ces spécificités autoriseront ainsi l’étude des phénomènes d’induction de tumeurs du système nerveux à l’échelle de la cellule individuelle, ce qui est impossible dans les modèles vertébrés du fait de l’extrême complexité du cerveau. Ce modèle d’étude particulièrement prometteur devrait nous permettre de comprendre comment une cellule nerveuse peut acquérir des caractéristiques tumorales pendant l’embryogenèse et de percer le mystère des mécanismes qui conduisent à l’initiation de tumeurs pédiatriques du cerveau.
II/ Projets de recherche translationnelle
II/1) Ciblage de la différentiation tumorale dans les médulloblastomes et les glioblastomes
Les thérapies actuelles impliquent des protocoles de chimiothérapie et de radiothérapie qui ciblent exclusivement les cellules en division au sein de la masse tumorale. Ces traitements ne permettent pas toujours l’éradication de toutes les cellules cancéreuses. En effet, certaines sous-populations de cellules, présentant notamment une prolifération lente voire quiescentes et des caractéristiques de cellules souches semblent plus résistantes aux traitements génotoxiques. Ainsi, ces cellules peuvent persister et engendrer des récidives. Le traitement des tumeurs cérébrales par irradiation apparaît actuellement comme étant le plus efficace des traitements non chirurgicaux. Cependant, l’existence de cellules tumorales résistantes et leur enrichissement au sein de la tumeur après irradiation limite l’efficacité de celle-ci. Ces cellules présentant des propriétés souches, nous faisons l’hypothèse que le fait d’influer sur leur identité en les engageant vers une voie de différentiation permettrait d’augmenter la réponse anti-tumorale. Faisant suite au travail de recherche fondamentale (voir partie « EmbryOnco »), des gènes influençant l’état souche des cellules ont été mis en évidence par l’équipe. L’utilisation d’inhibiteurs de ces derniers en combinaison à une radiothérapie permettrait de sensibiliser les cellules tumorales à la radiothérapie et donc d’éviter les récidives (Filipponi et al. 2022).
II/2) Thérapies innovantes en médecine nucléaire
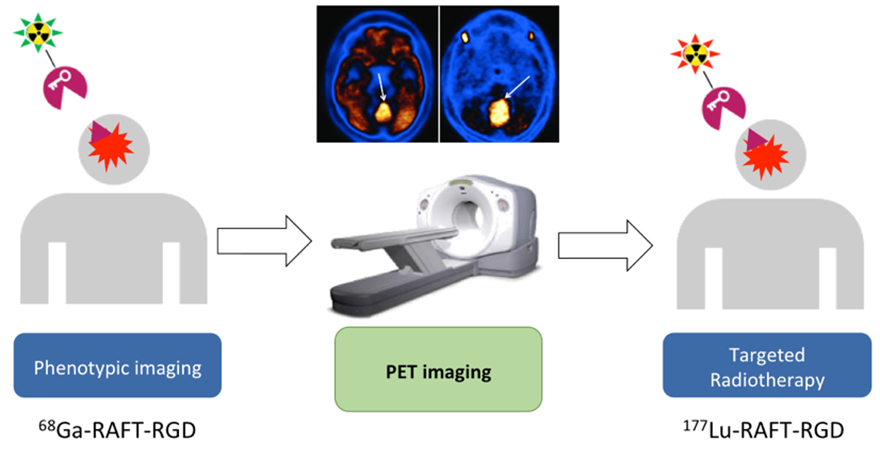
L’un des défis majeurs dans la prise en charge des tumeurs cérébrales pédiatriques ou adultes repose sur la nécessité de limiter au maximum les effets indésirables liés aux traitements. Une approche en plein essor en médecine nucléaire est l’approche thérapeutique théranostique. Elle consiste en une combinaison de diagnostic par imagerie nucléaire et de radiothérapie ; le théranostique utilise des composés qui permettent à la fois une détection spécifique des cellules cancéreuses et un traitement ciblé de la tumeur. Ces composés de type "missile à tête chercheuse" sont conçus pour reconnaître spécifiquement les marqueurs des cellules cancéreuses et détruire ensuite ces cellules. Les récentes avancées dans le domaine du théranostique ont permis des progrès remarquables dans la prise en charge du cancer de la prostate et des tumeurs neuroendocrines, tout en présentant des effets secondaires limités.
Le développement d’une telle approche nécessite une cible d’intérêt et un radiotraceur pertinent. De nombreux travaux ont en lumière le caractère essentiel des interactions matrice extracellulaire-cellules tumorales dans la tumorigénèse des tumeurs cérébrales (caractère souche, invasion, angiogenèse). Dans l’ensemble de ces mécanismes, les intégrines, et plus particulièrement l’intégrine αvβ3 jouent un rôle fondamental.
Ces données ont conduit au développement subséquent de nombreux agents thérapeutiques, basés sur l’utilisation de dérivés du RGD (Arg-Gly-Asp), motif liant αvβ3, dont le cilengitide. L’équipe a récemment démontré que l’intégrine αvβ3 est un acteur clé de la tumorigenèse des médulloblastomes. L’équipe collabore, pour ce projet avec le Laboratoire Radiopharmaceutiques Biocliniques (Grenoble, France), spécialiste du développement d’agents théranostiques et du ciblage théranostique de l’intégrine αvβ3.
Le LRB a précédemment développé un ligand de l'intégrine αvβ3, le RAFT-RGD. Le radiomarquage au 68Ga- pour l’imagerie pré-thérapeutique et au 177Lu- pour la radiothérapie interne vectorisée.
Le développement pré-clinique de ce projet est étroitement lié au prochain transfert clinique de la molécule pour la prise en charge des glioblastomes.
II/3) Radiothérapie et ferroptose (Ferrox-RT : Ferroptosis and Oxydative Stress in Radiotherapy)
Augmenter l’efficacité de la radiothérapie et limiter ses effets secondaires font partie des enjeux majeurs dans la prise en charge des tumeurs cérébrales. De récents travaux, et notamment ceux menées par l’équipe « Hypoxie tumorale et métabolisme » au CSM, ont mis en lumière une nouvelle forme de mort cellulaire : la ferroptose. Celle-ci résulte d’une accumulation de lipides peroxydés au niveau des membranes cellulaires. Cette mort cellulaire est dépendante du fer et se déroule lors d’un stress oxydant important : le fer libre permet la transformation d’espèces oxydantes faiblement actives (H2O2) en des oxydants très puissants, comme le radical hydroxyle (OH) capables d’oxyder les lipides insaturés, déclenchant une réaction en chaîne qui peut aboutir à la mort des cellules.
Les rayonnements ionisants utilisés pour la radiothérapie provoquent la radiolyse l’eau intracellulaire, ce qui a pour effet de libérer des ROS. Ainsi, la radiothérapie peut être à l’origine de l’induction d’un stress oxydant très important qui peut conduire à l’activation des voies de ferroptose. Inversement, une forte résistance de certaines cellules à la ferroptose peut se traduire par une radiorésistance et une mauvaise réponse à la radiothérapie. Comprendre les phénomènes de ferroptose radio-induite permettra ainsi de mieux comprendre certains mécanismes de radiorésistance mis en jeu dans les tumeurs présentant une faible réponse à la radiothérapie.
Par ailleurs, des études ont mis en évidence l’importance des interactions MEC-cellule tumorale dans la résistance à la mort par ferroptose notamment via l’expression de certaines intégrines. Nos études préliminaires ont démontré que l’intégrine αvβ3 est étroitement associé à une résistance à la radiothérapie et à une résistance vis à vis de la mort par ferroptose.
L’objectif de ce projet est à la fois très fondamental et translationnel puisqu’il vise à (i) décrypter le rôle de l’intégrine αvβ3 dans la résistance à la radiothérapie et à la ferroptose et (ii) proposer une radiosensibilisation des tumeurs cérébrales en ciblant les régulateurs de ces mécanismes.
Bibliographie
1. Luo, Z., Xia, M., Shi, W., Zhao, C., Wang, J., Xin, D., Dong, X., Xiong, Y., Zhang, F., Berry, K., et al. (2022). Human fetal cerebellar cell atlas informs medulloblastoma origin and oncogenesis. Nature, 1–8. 10.1038/s41586-022-05487-2.
2. Smith, K.S., Bihannic, L., Gudenas, B.L., Haldipur, P., Tao, R., Gao, Q., Li, Y., Aldinger, K.A., Iskusnykh, I.Y., Chizhikov, V.V., et al. (2022). Unified rhombic lip origins of group 3 and group 4 medulloblastoma. Nature 609, 1012–1020. 10.1038/s41586-022-05208-9.
3. Zhao, S., Li, J., Zhang, H., Qi, L., Du, Y., Kogiso, M., Braun, F.K., Xiao, S., Huang, Y., Li, J., et al. (2022). Epigenetic Alterations of Repeated Relapses in Patient-matched Childhood Ependymomas. Nat. Commun. 13, 6689. 10.1038/s41467-022-34514-z.
4. Minniti, G., Niyazi, M., Alongi, F., Navarria, P., and Belka, C. (2021). Current status and recent advances in reirradiation of glioblastoma. Radiat. Oncol. Lond. Engl. 16, 36. 10.1186/s13014-021-01767-9.
5. Kumar, R., Smith, K.S., Deng, M., Terhune, C., Robinson, G.W., Orr, B.A., Liu, A.P.Y., Lin, T., Billups, C.A., Chintagumpala, M., et al. (2021). Clinical Outcomes and Patient-Matched Molecular Composition of Relapsed Medulloblastoma. J. Clin. Oncol. 39, 807–821. 10.1200/JCO.20.01359.
6. Salloum, R., Chen, Y., Yasui, Y., Packer, R., Leisenring, W., Wells, E., King, A., Howell, R., Gibson, T.M., Krull, K.R., et al. (2019). Late Morbidity and Mortality Among Medulloblastoma Survivors Diagnosed Across Three Decades: A Report From the Childhood Cancer Survivor Study. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 37, 731–740. 10.1200/JCO.18.00969.
7. Mannino, M., and Chalmers, A.J. (2011). Radioresistance of glioma stem cells: Intrinsic characteristic or property of the ‘microenvironment‐stem cell unit’? Mol. Oncol. 5, 374–386. 10.1016/j.molonc.2011.05.001.
Publications
-
A fistful of iron: ferritin as a vulnerability point of the brain cancers
Segui F, Parks SK, Vucetic M, Picco V
Cell Death Dis -
Development and validation of a PSMA-positive triple-negative breast cancer mouse model for preclinical targeted radionuclide therapies
Chaussin B, Sanchez L, Levesque S, Revy A, Tempier M, Montemagno C, Durivault J, Schmitt S, Boutault E, Besse S, Chautard E, Auriol M, Voissiere A, Kossaï M, Miot-Noirault E, Penault-Llorca F, Merlin C, Cachin F, Jouberton E
Sci Rep -
Feasibility of In Vivo Dosimetry in Contact Radiotherapy Using Germanium-Doped Silica Optical Fibers
Shumilov G, Gentner C, Amblard R, Villeneuve R, Garnier N, Benezery K, Montemagno C, Picco V, El Hamzaoui H, Bouet M, Capoen B, Bouazaoui M, Mady F, Benabdesselam M, Serrano B
Phys. Status Solidi A Volume : 223, Issue : 2, e202500633 -
Genomic determinants of Bacillus cereus and outcomes of infection in preterm neonates: a multicentre retrospective study
Chevalier A, Ben Khedher M, Tran A, Coin M, Pousse M, Tazi A, Eleni Dit Trolli S, Lanotte P, Follet C, Pichon M, Dubois D, Herbulot L, Potron A, Aberrane S, Birgy A, Boutillier B, Pantel A, Di Maio M, Dubourg G, Boyer P, Corvec S, Groshenry G, Bonis M, Saissi M, Gallet A, Boyer L, Croce O, Lotte R, Ruimy R
Clinical Microbiology and Infection -
Neural Innervation of Tumors: Mechanisms, Hallmarks, and Therapeutic Opportunities
Cassim S, Montemagno C
Cancers Volume 18, Issue 7, Page 1063
-
Comparative theranostic efficacy of 177Lu- and 161Tb-labeled A1K2 SdAb in mesothelin-positive tumors
N’Guessan E, Raes F, Ahmadi M, Bacot S, Dumas L, Leenhardt J, Du Moulinet d’Hardemare A, Debiossat M, André C, Boutonnat J, Durivault J, Montemagno C, Lenormand JL, Djaïleb L, Köster U, Van de Voorde M, Ramaekers S, Verguts K, Ghezzi C, Perret P, Lombardi C, Broisat A
Eur J Nucl Med Mol Imaging -
De Novo Serine Synthesis Is a Metabolic Vulnerability That Can Be Exploited to Overcome Sunitinib Resistance in Advanced Renal Cell Carcinoma
Teisseire M, Sahu U, Parola J, Tsai M-C, Vial V, Durivault J, Grépin R, Cormerais Y, Molina C, Gouraud A, Pagès G, Ben-Sahra I, Giuliano S
Cancer Res 85 (10): 1857–1873 -
From Seeing to Healing: The Clinical Potential of Radiotracers in Pediatric Neuro-Oncology
Bogdanovic B, Montemagno C
Cancers 17(12):1905 -
Targeting Melanin Heterogeneity in Metastatic Melanoma: A Dual-Tumour Mouse Melanoma Model
Delmas M, Chaussin B, Harismendy N, Dougé A, Rouzaire P-O, Montemagno C, Durivault J, Moreau E, Miot-Noirault E, Quintana M, Besse S, D'Incan M, Chautard E, Jouberton E, Rouanet J
Exp Dermatol 34, no. 9 (2025): e70159 -
Theranostics in Hematological Malignancies: Cutting-Edge Advances in Diagnosis and Targeted Therapy
Bogdanovic B, Hugonnet F, Montemagno C
Cancers 17(7), 1247 -
Unveiling the Microeukaryotic Landscape of the Red Coral Corallium rubrum Across the Northwestern Mediterranean Sea
Prioux C, Filipponi D, Ferrier-Pagès C, Allemand D, Tignat-Perrier R
Environ Microbiol Volume 17, Issue 6, e70227
-
Exploiting Integrin-αVβ3 to Enhance Radiotherapy Efficacy in Medulloblastoma via Ferroptosis
Gotorbe C, Segui F, Echavidre W, Durivault J, Blanchard T, Vial V, Pagnuzzi-Boncompagni M, Villeneuve R, Amblard R, Garnier N, Ortholan C, Serrano B, Picco V, Pouyssegur J, Vucetic M, Montemagno C
Curr. Oncol. 31, 7390–7402 -
Integrin Targeting and Beyond: Enhancing Cancer Treatment with Dual-Targeting RGD (Arginine–Glycine–Aspartate) Strategies
Bogdanovic B, Fagret D, Ghezzi C, Montemagno C
Pharmaceuticals 17(11), 1556 -
Revealing the Ferroptotic Phenotype of Medulloblastoma
Segui F, Daher B, Gotorbe C, Pouyssegur J, Picco V, Vucetic M
J Vis Exp 2024 Mar 15:(205) -
Sulforaphane prevents diabetes-induced hepatic ferroptosis by activating Nrf2 signaling axis
Savic N, Markelic M, Stancic A, Velickovic K, Grigorov I, Vucetic M, Martinovic V, Gudelj A, Otasevic V
Biofactors 50(4):810-827 -
Unveiling CXCR2 as a promising therapeutic target in renal cell carcinoma: exploring the immunotherapeutic paradigm shift through its inhibition by RCT001
Montemagno C, Jacquel A, Pandiani C, Rastoin O, Dawaliby R, Schmitt T, Bourgoin M, Palenzuela H, Rossi A-L, Ambrosetti D, Durivault J, Luciano F, Borchiellini D, Le Du J, Pires Gonçalves L C, Auberger P, Benhida R, Kinget L, Beuselinck B, Ronco C, Pagès G, Dufies M
J Exp Clinical Cancer Research 43(1):86
-
A group of novel VEGF splice variants as alternative therapeutic targets in renal cell carcinoma
Montemagno C, Durivault J, Gastaldi C, Dufies M, Vial V, He X, Ambrosetti D, Kamenskaya A, Négrier S, Bernhard J-C, Borchiellini D, Cao Y, Pagès G
mol 21 Feb 2023, 45 -
Integrin-αvβ3 is a Therapeutically Targetable Fundamental Factor in Medulloblastoma Tumorigenicity and Radioresistance
Echavidre W, Durivault J, Gotorbe C, Blanchard T, Pagnuzzi M, Vial V, Raes F, Broisat A, Villeneuve R, Amblard R, Garnier N, Ortholan C, Faraggi M, Serrano B, Picco V, Montemagno C
Cancer Res Commun 3(12):2483-2496 -
Quantitative analysis of 99mTc‐pertechnetate thyroid uptake with a large‐field CZT gamma camera: feasibility and comparison between SPECT/CT and planar acquisitions
Serrano B, Amblard R, Beaumont T, Hugonnet F, Dietz M, Berthier F, Garnier N, Villeneuve R, Nataf V, Mocquot F, Montemagno C, Faraggi M, Paulmier B
EJNMMI Phys 10, 45 (2023) -
Recent Pre-Clinical Advancements in Nuclear Medicine: Pioneering the Path to a Limitless Future
Echavidre W, Fagret D, Faraggi M, Picco V, Montemagno C
Cancers 15(19), 4839 -
Should evidence of an autolysosomal de-acidification defect in Alzheimer and Parkinson diseases call for caution in prescribing chronic PPI and DMARD?
Giuliano S, Montemagno C, Domdom M-A, Teisseire M, Brest P, Klionsky D J, Hofman P, Pagès G, Mograbi B
Autophagy 19(10):2800-2806
-
Antiangiogenic Compound Axitinib Demonstrates Low Toxicity and Antitumoral Effects against Medulloblastoma
Pagnuzzi M, Picco V, Vial V, Planas-Bielsa V, Vandenberghe A, Daubon T, Derieppe M-A, Montemagno C, Durivault J, Grépin R, Martial S, Doyen J, Gavard J, Pagès G
Cancers 14(1) : 70 -
Cancer-associated fibroblasts in renal cell carcinoma: implication in prognosis and resistance to anti- angiogenic therapy
Ambrosetti D, Coutts M, Paoli C, Durand M, Borchiellini D, Montemagno C, Rastoin O, Borderie A, Grépin R, Rioux-Leclercq N, Bernhard J-C, Pagès G, Dufies M
BJU Int 2022 Jan;129(1):80-92 -
Ferroptosis as a Novel Determinant of β-Cell Death in Diabetic Conditions
Stancic A, Saksida T, Markelic M, Vucetic M, Grigorov I, Martinovic V, Gajic D, Ivanovic A, Velickovic K, Savic N, Otasevic V
Oxidative Medicine and Cellular Longevity 2022:3873420 -
Inhibiting ALK2/ALK3 Signaling to Differentiate and Chemo-Sensitize Medulloblastoma
Filipponi, D, Pagnuzzi M, Pagès G
Cancers 14(9): 2095 -
Integrin-αvβ3 as a Therapeutic Target in Glioblastoma: Back to the Future?
Echavidre W, Picco V, Faraggi M, Montemagno C
Pharm. 14(5) : 1053 -
Opposing Roles of Vascular Endothelial Growth Factor C in Metastatic Dissemination and Resistance to Radio/Chemotherapy: Discussion of Mechanisms and Therapeutic Strategies
Montemagno C, Luciano F, Pagès G
Methods Mol Biol 2475:1-23 -
Targeting of c-MET and AXL by cabozantinib is a potential therapeutic strategy for patients with head and neck cell carcinoma
Hagege A, Saada-Bouzid E, Ambrosetti D, Rastoin O, Boyer J, He X, Rousset J, Montemagno C, Doyen J, Pedeutour F, Parola J, Bourget I, Luciano F, Bozec A, Cao Y, Pagès G, Dufies M
Cell Reports Medicine 3(9): 100659 -
Targeting of the ELR+CXCL/CXCR1/2 Pathway Is a Relevant Strategy for the Treatment of Paediatric Medulloblastomas
Penco-Campillo M, Molina C, Piris P, Soufi N, Carré M, Pagnuzzi M, Picco V, , Dufies M, Ronco C, Benhida R, Martial S, Pagès G
Cells 11(23), 3933
-
Anti-Vascular Endothelial Growth Factor C Antibodies Efficiently Inhibit the Growth of Experimental Clear Cell Renal Cell Carcinomas
Dumond A, Montemagno C, Vial V, Grépin R, Pagès G
Cells 10(5): 1222 -
Cancer associated fibroblasts in renal cell carcinoma: implication in prognosis and resistance to antiangiogenic therapy
Ambrosetti D, Coutts M, Paoli C, Durand M, Borchiellini D, Montemagno C, Rastoin O, Borderie A, Grépin R, Rioux-Leclercq N, Bernhard J-C, Pagès G, Dufies M
BJU Int doi: 10.1111/bju.15506. -
Experimental and computational modeling for signature and biomarker discovery of renal cell carcinoma progression
Cooley L S, Rudewicz J, Souleyreau W, Emanuelli A, Alvarez-Arenas A, Clarke K, Falciani F, Dufies M, Lambrechts D, Modave E, Chalopin-Fillot D, Pineau R, Ambrosetti D, Bernhard J-C, Ravaud A, Négrier S, Ferrero J-M, Pagès G, Benzekry S, Nikolsky M, Bikfalvi A
Mol. Cancer 20:136 -
Hypoxia and hypoxia-inducible factors promote the development of neointimal hyperplasia in arteriovenous fistula
Sadaghianloo N, Contenti J, Declemy S, Ambrosetti D, Zdralevic M, Tannour-Louet M, Fabbri L, Pagès G, Bost F, Hassen-Khodja R, Pouyssegur J, Jean-Baptise E, Dardik A, Mazure N M
J Physiol 599: 2299-2321 -
In vivo monitoring of the therapeutic efficacy of a CXCR1/2 inhibitor with 18F-FDG PET/CT imaging in experimental head and neck carcinoma: A feasibility study
Montemagno C, Serrano B, Durivault J, Nataf V, Mocquot F, Amblard R, Vial V, Ronco C, Benhida R, Dufies M, Faraggi M, Pagès G
Biochem Biophys Rep 27: 101098 -
Neuropilin 1 and Neuropilin 2 gene invalidation or pharmacological inhibition reveals their relevance for the treatment of metastatic renal cell carcinoma
Dumond A, Brachet E, Durivault J, Vial V, Puszko A K, Lepelletier Y, Montemagno C, Pagnuzzi M, Hermine O, Garbay C, Lagarde N, Montes M, Demange L, Grépin R, Pagès G
J Exp Clinical Cancer Research 40: 33 -
Pancreatic Ductal Adenocarcinoma: The Dawn of the Era of Nuclear Medicine?
Montemagno C, Cassim S, De Leiris N, Durivault J, Faraggi M, Pagès G
Int J Mol Sci 22, 6413 -
Plk1, upregulated by HIF-2, mediates metastasis and drug resistance of clear cell renal cell carcinoma
Dufies M, Verbiest A, Cooley L S, Ndiaye P D, He W, Nottet N, Souleyreau W, Hagege A, Torrino S, Parola J, Giuliano S, Borchiellini D, Schiappa R, Mograbi B, Zucman-Rossi J, Bensalah K, Ravaud A, Auberger P, Bikfalvi A, Chamorey E, Rioux-Leclercq N, Mazure N M, Beuselinck B, Cao Y, Bernhard J C, Ambrosetti D, Pagès G
Comm Biol 4: 166 -
Relevance of Neuropilin 1 and Neuropilin 2 targeting for cancer treatment
Dumond A, Pagès G
J Cancer Immunol 3(2): 111-114 -
The Polo-like kinase 1 inhibitor onvansertib represents a relevant treatment for head and neck squamous cell carcinoma resistant to cisplatin and radiotherapy
Hagege A, Ambrosetti D, Boyer J, Bozec A, Doyen J, Chamorey E, He X, Bourget I, Rousset J, Saada E, Rastoin O, Parola J, Luciano F, Cao Y, Pagès G, Dufies M
Theranostics 11(19): 9571-9586 -
UBTD1 regulates ceramide balance and endolysosomal positioning to coordinate EGFR signaling
Torrino S, Tiroille V, Dolfi B, Dufies M, Hinault C, Bonesso L, Dagnino S, Uhler J, Irondelle M, Gay A S, Fleuriot L, Debayle D, Lacas-Gervais S, Cormont M, Bertero T, Bost F, Gilleron J, Clavel S
Elife 10(e68348): 1-24
-
Co-culture of human fibroblasts, smooth muscle and endothelial cells promotes osteopontin induction in hypoxia
Sadaghianloo N, Contenti J, Dufies M, Parola J, Rouleau M, Lee S, Peyron J-F, Fabbri L, Hassen-Khodja R, Pouyssegur J, Bost F, Jean-Baptiste E, Dardik A, Mazure N M
J Cell Mol Med 24(5): 2931-2941 -
Depicting the genetic architecture of pediatric cancers through an integrative gene network approach
Savary C, Kim A, Lespagnol A, Gandemer V, Pellier I, Andrieu C, Pagès G, Galibert M-D, Blum Y, de Tayrac M
Sci Rep 10: 1224 -
Evidences of a direct relationship between cellular fuel supply and ciliogenesis regulated by hypoxic VDAC1-∆C
Meyenberg Cunha-de Padua M, Fabbri L, Dufies M, Lacas-Gervais S, Contenti J, Voyton C, Fazio S, Irondelle M, Mograbi B, Rouleau M, Sadaghianloo N, Rovini A, Brenner C, Craigen W J, Bourgeais J, Herault O, Bost F, Mazure N M
Cancers 12(11): 3484 -
Experimental models in neovascular age related macular degeneration
Rastoin O, Pagès G, Dufies M
Int J Mol Sci 21: 4627 -
From Malignant Progression to Therapeutic Targeting: Current Insights of Mesothelin in Pancreatic Ductal Adenocarcinoma
Montemagno C, Cassim S, Pouyssegur J, Broisat A, Pagès G
Int J Mol Sci 21(11): 4067 -
Identification of a new aggressive axis driven by ciliogenesis and absence of VDAC1-ΔC in clear cell Renal Cell Carcinoma patients
Dufies M, Fabbri L, Lacas-Gervais S, Gardie B, Gad-Lapiteau S, Parola J, Nottet N, Meyenberg Cunha de Padua M, Contenti J, Borchiellini D, Ferrero J-M, Rioux Leclercq N, Ambrosetti D, Mograbi B, Richard S, Viotti J, Chamorey E, Sadaghianloo N, Rouleau M, Craigen W J, Mari B, Clavel S, Pagès G, Pouyssegur J, Bost F, Mazure N M
Theranostics 10(6): 2696-2713 -
Les neuropilines : Des cibles pertinentes pour améliorer le traitement des cancers
Dumond A, Demange L, Pagès G
Med Sci (Paris) 36(5): 487-496 -
Metastatic Heterogeneity of Breast Cancer: Companion and Theranostic Approach in Nuclear Medicine
Montemagno C, Pagès G
Cancers 12(4): 821 -
Neuropilins, as Relevant Oncology Target: Their Role in the Tumoral Microenvironment
Dumond A, Pagès G
Front Cell Dev Biol 8: 662 -
Resistance to anti-angiogenic therapies: A mechanism depending on the time of exposure to the drugs
Montemagno C, Pagès G
Front Cell Dev Biol 8: 584 -
SH3BGRL2, a new down regulated tumor suppressor in clear cell renal cell carcinomas
Dufies M
EBioMedicine 52: 102641 -
Soluble forms of PD-L1 and PD-1 as prognostic and predictive markers of sunitinib efficacy in patients with metastatic clear cell renal cell carcinoma.
Montemagno C, Hagege A, Borchiellini D, Thamphya B, Rastoin O, Ambrosetti D, Iovanna J, Rioux-Leclercq N, Porta C, Négrier S, Ferrero J-M, Chamorey E, Pagès G, Dufies M
Oncoimmunology 9:1, 1846901 -
Synthesis and biological evaluation of 3-amino-1,2,4-triazole derivatives as potential anticancer compounds
Grytsai O, Valiashko O, Penco-Campillo M, Dufies M, Hagege A, Demange L, Martial S, Pagès G, Ronco C, Benhida R
Bioorg Chem 104: 104271 -
Targeting mesothelin in pancreatic ductal adeno-carcinoma (PDAC)
Montemagno C, Pagès G
J Cell Immunol 2(2): 65-67 -
The combination of bevacizumab/Avastin and erlotinib/Tarceva is relevant for the treatment of metastatic renal cell carcinoma: the role of a synonymous mutation of the EGFR receptor
Grépin R, Guyot M, Dumond A, Durivault J, Ambrosetti D, Roussel J F, Dupré F, Quintens H, Pagès G
Theranostics 10(3): 1107-1121 -
VEGFC negatively regulates the growth and aggressiveness of medulloblastoma cells
Penco-Campillo M, Comoglio Y, Feliz Morel A J, Hanna R, Durivault J, Leloire M, Mejias B, Pagnuzzi M, Morot A, Burel-Vandenbos F, Selby M, Williamson D, Clifford S C, Claren A, Doyen J, Picco V, Martial S, Pagès G
Comm Biol 3(758): 579
-
99mTc-A1 as a Novel Imaging Agent Targeting Mesothelin-Expressing Pancreatic Ductal Adenocarcinoma
Montemagno C, Cassim S, Trichanh D, Savary C, Pouyssegur J, Pagès G, Fagret D, Broisat A, Ghezzi C
Cancers 11(10): 1531 -
In Vivo Assessment of VCAM-1 Expression by SPECT/CT Imaging in Mice Models of Human Triple Negative Breast Cancer.
Montemagno C , Dumas L, Cavaillès P, Ahmadi M, Bacot S, Debiossat M, Soubies A, Djaïleb L, Leenhardt J, De Leiris N, Dufies M, Pagès G, Hernot S, Devoogdt N, Perret P, Riou L, Fagret D, Ghezzi C, Broisat A
Cancers 11(7). pii: E1039 -
Inhibition of the amino‐acid transporter LAT1 demonstrates anti‐neoplastic activity in medulloblastoma
Cormerais Y, Pagnuzzi M, Schrötter S, Giuliano S, Tambutté E, Endou H, Wempe M F, Pagès G, Pouyssegur J, Picco V
J Cell Mol Med 1-8 -
Large expert-curated database for benchmarking document similarity detection in biomedical literature search
Brown P, Relish Consortium, Dufies M, Zhou Y
Database baz085 -
New CXCR1/CXCR2 inhibitors represent an effective treatment for kidney or head and neck cancers sensitive or refractory to reference treatments
Dufies M, Grytsai O, Ronco C, Camara O, Ambrosetti D, Hagege A, Parola J, Mateo L, Ayrault M, Giuliano S, Grépin R, Lagarde N, Montes M, Auberger P, Demange L, Benhida R, Pagès G
Theranostics 9(18): 5332–5346 -
PGC-1α inhibits polyamine synthesis to suppress prostate cancer aggressiveness
Kaminski L, Torrino S, Dufies M, Djabari Z, Haider R, Roustan F-R, Jaune E, Laurent K, Nottet N, Michiels J-F, Gesson M, Rocchi S, Mazure N M, Durand M, Tanti J-F, Ambrosetti D, Clavel S, Ben-Sahra I, Bost F
Cancer Res 79(13): 3268-3280 -
Resistance to lysosomotropic drugs used to treat kidney and breast cancers involves autophagy and inflammation and converges in inducing CXCL5
Giuliano S, Dufies M, Ndiaye P D, Viotti J, Borchiellini D, Parola J, Vial V, Cormerais Y, Ohanna M, Imbert V, Chamorey E, Rioux-Leclercq N, Savina A, Ferrero J-M, Mograbi B, Pagès G
Theranostics 9(4): 1181-1199 -
Synthesis, 3D-structure and stability analyses of NRPa-308, a new promising anti-cancer agent
Brachet E, Dumond A, Liu W-Q, Fabre M, Selkti M, Raynaud F, Hermine O, Benhida R, Belmont P, Garbay C, Lepelletier Y, Ronco C, Pagès G, Demange L
Bioorg Med Chem Lett 29:126710 -
VEGF-C et vaisseaux lymphatiques. Une épée à double tranchant dans le développement tumoral et la dissémination métastatique
Ndiaye P D, Pagès G
Med Sci (Paris) 2(35): 132-137 -
VEGFC acts as a double-edged sword in renal cell carcinoma aggressiveness
Ndiaye P D, Dufies M, Giuliano S, Douguet L, Grépin R, Durivault J, Lenormand P, Glisse N, Mintcheva J, Vouret-Craviari V, Mograbi B, Wurmser M, Ambrosetti D, Rioux-Leclercq N, Maire P, Pagès G
Theranostics 9(3): 661-675
-
AKT1 restricts the invasive capacity of head and neck carcinoma cells harboring a constitutively active PI3 kinase activity
Brolih S, Parks S, Vial V, Durivault J, Mostosi L, Pouyssegur J, Pagès G, Picco V
BMC Cancer 18(1): 249 -
Soluble CD146 is a predictive marker of pejorative evolution and of sunitinib efficacy in clear cell renal cell carcinoma
Dufies M, Nollet M, Ambrosetti D, Traboulsi W, Viotti J, Borchiellini D, Grépin R, Parola J, Giuliano S, Helley-Russick D, Bensalah K, Ravaud A, Bernhard J C, Schiappa R, Bardin N, Dignat-George F, Rioux-Leclercq N, Oudard S, Négrier S, Ferrero J M, Chamorey E, Blot-Chabaud M, Pagès G
Theranostics 8(9): 2447-2458 -
The path to « the Golden Age » for the treatment of metastatic renal cell carcinoma
Pagès G
Oncotarget 9(60): 31564-31565 -
The two glycolytic markers GLUT1 and MCT1 correlate with tumor grade and survival in clear-cell renal cell carcinoma
Ambrosetti D, Dufies M, Dadone B, Durand M, Borchiellini D, Amiel J, Pouyssegur J, Rioux-Leclercq N, Pagès G, Burel-Vandenbos F, Mazure N
PLOS O 13(2): e0193477 -
The tyrosine kinase inhibitor Sunitinib targets Vascular Endothelial (VE)-cadherin: a marker of response to antitumoral treatment in metastatic renal cell carcinoma
Polena H, Creuzet J, Dufies M, Sidibé A, Khalil-Mgharbel A, Salomon A, Deroux A, Quesada J-L, Roelants C, Filhol O, Cochet C, Blanc E, Ferlay-Segura C, Borchiellini D, Ferrero J-M, Escudier B, Négrier S, Pagès G, Vilgrain I
Br J Cancer 118(9): 1179-1188
-
CXCL7 is a predictive marker of sunitinib efficacy in clear cell renal cell carcinomas
Dufies M, Giuliano S, Viotti J, Borchiellini D, Cooley L S, Ambrosetti D, Guyot M, Ndiaye P D, Parola J, Claren A, Schiappa R, Gal J, Frangeul A, Jacques A, Cassuto O, Grépin R, Auberger P, Bikfalvi A, Milano G, Escudier B, Rioux-Leclercq N, Porta C, Negrier S, Chamorey E, Ferrero J M, Pagès G
Br J Cancer 117(7): 947-953 -
NRPa-308, a new neuropilin-1 antagonist, exerts in vitro anti-angiogenic and anti-proliferative effects and in vivo anti-cancer effects in a mouse xenograft model
Liu W-Q, Lepelletier Y, Montes M, Borriello L, Jarray R, Grépin R, Leforban B, Loukaci A, Benhida R, Hermine O, Dufour S, Pagès G, Garbay C, Raynaud F, Hadj-Slimane R, Demange L
Can Let 414: 88-98 -
Structural and functional analysis of coral Hypoxia Inducible Factor
Zoccola D, Morain J, Pagès G, Caminiti Segonds N, Giuliano S, Tambutté S, Allemand D
PLOS O 12(11): e0186262 -
Sunitinib Stimulates Expression of VEGFC by Tumor Cells and Promotes Lymphangiogenesis in Clear Cell Renal Cell Carcinomas.
Dufies M, Giuliano S, Ambrosetti D, Claren A, Ndiaye P D, Mastri M, Moghrabi W, Cooley L S, Ettaiche M, Chamorey E, Parola J, Vial V, Lupus-Plesu M, Bernhard J C, Ravaud A, Borchiellini D, Ferrero J M, Bikfalvi A, Ebos J M, Khabar K S, Grépin R, Pagès G
Cancer Res 77(5): 1-15 -
Targeting the pro-angiogenic forms of VEGF or inhibiting their expression as anti-cancer strategies
Guyot M, Hilmi C, Ambrosetti D, Merlano M, Lo Nigro C, Durivault J, Grépin R, Pagès G
Oncotarget 8(6): 9174-9188 -
The c-Jun N-terminal kinase prevents oxidative stress induced by UV and thermal stresses in corals and human cells
Picco V, Courtial L, Grover R, Cormerais Y, Rottier C, Labbe A, Pagès G, Ferrier-Pagès C
Sci Rep 7: 45713 -
Validation of commercial ERK antibodies against the ERK orthologue of the scleractinian coral Stylophora pistillata
Picco V, Courtial L, Pagès G, Ferrier-Pagès C
F1000Res 6(577)
-
ERK1/2/MAPK pathway-dependent regulation of the telomeric factor TRF2
Picco V, Coste I, Giraud-Panis M-J, Renno T, Gilson E, Pagès G
Oncotarget 7(29): 46615-46627 -
Lebein, a snake venom disintegrin, suppresses human colon cancer cells proliferation and tumor-induced angiogenesis through cell cycle arrest, apoptosis induction and inhibition of VEGF expression.
Zakraoui O, Marcinkiewicz C, Aloui Z, Othman H, Grépin R, Haoues M, Essafi M, Srairi-Abid N, Gasmi A, Karoui H, Pagès G, Essafi-Benkhadir K
Mol Carcinog 56(1): 18-35 -
Systematic analysis of AU-rich element expression in cancer reveals common functional clusters regulated by key RNA-binding proteins.
Hitti E, Bakheet T, Al-Souhibani N, Moghrabi W, Al-Yahya S, Al-Ghamdi M, Al-Saif M, Shoukri M M, Lanczk A, Grépin R, Gyorffy B, Pagès G, Khabar K S
Cancer Res 76(14): 4068-4080 -
Telomeric repeat-binding factor 2: a marker for survival and anti-EGFR efficacy in oral carcinoma
Picco V, Benhamou Y, Raybaud H, Sudaka A, Chamorey E, Brolih S, Monteverde M, Merlano M, Nigro C L, Ambrosetti D, Pagès G
Oncotarget 7(28): 44236-44251 -
The telomere proteins in tumorigenesis and clinical outcomes of oral squamous cell carcinoma
Benhamou Y, Picco V, Pagès G
Adaptation and Evolution in Marine Environments 57: 46-53
-
Resistance to Antiangiogenic Treatments via Upregulation of Substitution Pathways
Grépin R, Guyot M, Pagès G
Molecular Mechanisms of Angiogenesis Springer Paris, Chap. 20: 397-419 -
The CXCL7/CXCR1/2 axis is a key driver in the growth of clear cell renal cell carcinoma
Grépin R, Guyot M, Giuliano S, Pagnuzzi M, Ambrosetti D, Chamorey E, Scoazec J Y, Negrier S, Simonnet H, Pagès G
Cancer Res 74(3): 873-883 -
The ELRCXCL chemokines and their receptors CXCR1/CXCR2: A signaling axis and new target for the treatment of renal cell carcinoma
Giuliano S, Guyot M, Grépin R, Pagès G
Oncoimmunology 3: e28399 -
The Relevance of Testing the Efficacy of Anti-Angiogenesis Treatments on Cells Derived from Primary Tumors: A New Method for the Personalized Treatment of Renal Cell Carcinoma
Grépin R, Ambrosetti D, Marsaud A, Gastaud L, Amiel J, Pedeutour F, Pagès G
PLOS O 9(3): e89449